Dans la foule de messages alarmants sur le blanchiment du corail, suite au réchauffement climatique partout dans le monde, les résultats sur la résistance de certaines espèces coralliennes du Golfe d’Eilat semblent rassurants. Ils montrent que la résilience des écosystèmes marins face à des changements climatiques sont le produit de leur histoire évolutive à travers les temps géologiques. Les espèces qui peuplent aujourd’hui le nord de la Mer Rouge sont les mêmes qui se trouvent dans la région indopacifique. Ils ont donc dû migrer depuis l’Océan Indien par le sud de la Mer Rouge – aujourd’hui très chaud, mais encore plus adapté aux coraux il y a 18’000 ans, lors de la dernière glaciation. Ces espèces ont ensuite migré vers le nord en suivant le réchauffement postglaciaire. Au cours de ce voyage, seules les populations résistantes à des changements de température et de salinité ont survécu et sont arrivées au Golfe d’Eilat. Ce mécanisme de sélection naturelle explique leur tolérance aux expériences de surchauffe.
Cependant, une histoire qui a duré au moins 10’000 ans ne peut pas se faire en 150 ans. 150 ans c’est grosso modo le temps que l’Humanité a pris pour faire doubler le volume de CO2 dans l’atmosphère (jusqu’à 400 ppm) par rapport à la variation entre périodes glaciaires (180 ppm) et interglaciaires (280 ppm). C’est pourquoi les écosystèmes récifaux sensibles ne peuvent pas s’adapter.
Cette sensibilité est toutefois relative à l’endroit et aux conditions écologiques vécues par les populations durant les derniers millénaires, voire les derniers millions d’années. De plus, la hausse des températures n’est pas fatale à elle seule ; elle se combine en un cocktail de changements dont les plus significatifs sont l’eutrophisation (augmentation des nutriments) et l’acidification des mers qui peuvent rendre la calcification des organismes plus difficile.


Pour l’eutrophisation, regardons le cas des Caraïbes
Cette mer est parmi les régions les plus durement touchées par le dépérissement des récifs coralliens. Des espèces constructrices de barrières sont en voie de disparition (p. ex. Acropora cervicornis et A. palmata, voir figures 1 et 2).
De par son histoire, cette mer est déjà plus pauvre que d’autres en diversité corallienne, car elle est coupée du pool génétique de l’Indopacifique depuis la fermeture de passage marin en Amérique Centrale il y a 3 millions d’années environ. Il s’agit de populations avec peu de diversité génétique, donc plus vulnérables. Le corail de pierre des mers chaudes et lumineuses vit en symbiose avec des micro-algues qui produisent de la nourriture par photosynthèse à l’intérieur même du tissu corallien. C’est pourquoi ces récifs sont favorisés par des eaux claires, pauvres en nutriments et en plancton.
En 2005, le plus grand blanchiment du corail de l’histoire a été observé dans les Caraïbes et il a été attribué sans équivoque au réchauffement extrême (au delà de 30° pendant plusieurs semaines) de la partie sud-orientale (voir figures 3 et 4).
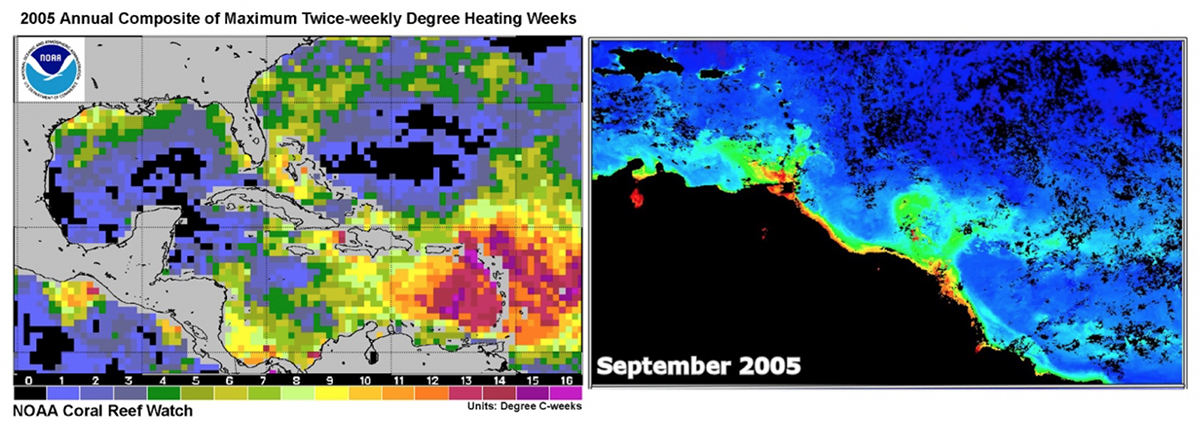
Fig. 4 Droite: Les panaches de l’Amazone et de l’Orénoque visualisés en quantités de couleur verte (chlorophylle et matière organique) colorée en septembre 2005 (maximum: rouge, minimum: bleu foncé). Image: NOAA.
En 2009-2010, notre équipe de recherche a entrepris une étude du plancton dans les Caraïbes sud-orientales (Cariacou, Barbade), une zone sous l’influence d’eaux de surface riches en nutriments qui sont produites par les panaches des fleuves Amazone et Orénoque et qui envahissent les Caraïbes orientales (voir figure). Lors des crues de ces 2 grands fleuves, nous avons observé un changement drastique dans la population corallienne vers des espèces planctoniques (radiolaires) opportunistes, typiques de milieux eutrophiques (très riches en nutriments).
Les panaches de l’Amazone et de l’Orénoque ont été étudiés récemment, car ils absorbent mieux le rayonnement solaire dans les couches de surface qui se réchauffent alors (elles sont d’ailleurs suspectées d’intensifier la violence des ouragans qui ont affecté les Caraïbes ces dernières décennies). Malgré les efforts consentis pour ces études, aucun lien entre les apports de nutriments et le dépérissement du corail n’a pu être établi.
Il existe pourtant une étroite corrélation entre la diminution des surfaces coralliennes en bonne santé et l’augmentation des surfaces déforestées des bassins versants des grands fleuves. Les panaches des fleuves sont un phénomène naturel et habituels ; par contre, leur teneur en nutriments, provenant de sols lessivés et ainsi artificiellement dépourvus de couverture végétale, augmente chaque année.
Ailleurs cependant, une expédition récente dans les récifs au large de Brunei (Bornéo) nous a révélé la relativité de ces phénomènes d’eutrophisation: on remarque sur la base de l’imagerie satellitaire que le niveau des nutriments dans les eaux de surface y est aussi importants que dans les Caraïbes sud-occidentales. Dans le sud de la Mer de Chine, la température estivale moyenne a augmenté entre 1950 et 2008 de 29° à 29.5° et les extrêmes mesurés au mois de novembre 2017 atteignaient 31°. Mais nous n’avons trouvé aucun blanchiment de corail, ni dans les atolls au grand large, ni dans des récifs côtiers à proximité d’estuaires. Malgré la présence abondante de plancton et par conséquent, des eaux assez peu transparentes, la couverture corallienne est intacte, très diverse et en bonne santé (voir figure 5 et 6). Comment expliquer ce phénomène ?


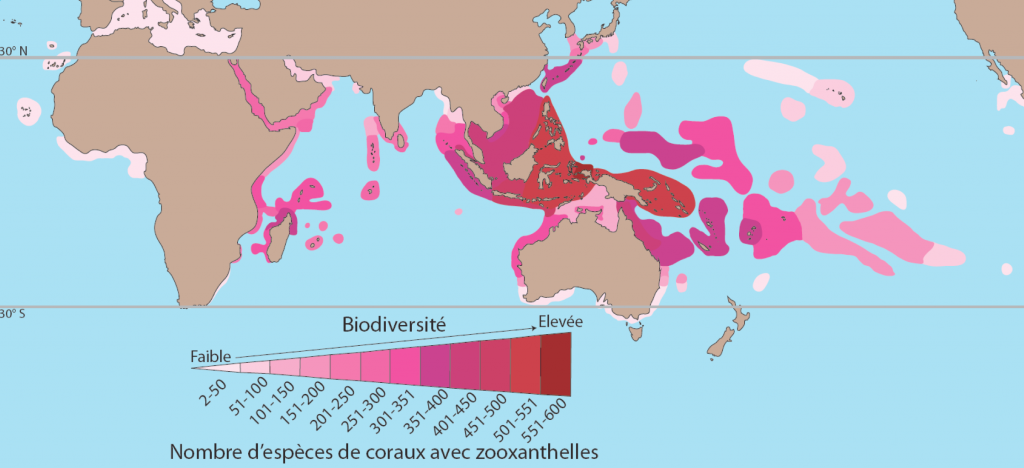
Encore une fois, c’est sur des périodes millénaires que les populations coralliennes ont pu évoluer dans un milieu plus riche en nutriments et plus chaud. En fait, la Mer de Chine fait partie du « Triangle de corail » (voir figure 7) où l’on a observé la plus grande diversité corallienne au monde – paradoxalement dans des zones assez riches en nutriments dérivés des terres tropicales pluvieuses.
Un autre phénomène d’adaptation de communautés récifales pendant des périodes millénaires sont les récifs découverts au large de l’embouchure de l’Amazone, découverts en 2016 sous les eaux troubles du panache. Peu étudiés, ils représentent des récifs eutrophiques, dominés par des espèces qui se nourrissent de la filtration du plancton (éponges, octocoralliens, etc.) plutôt que de la symbiose avec des algues comme les hexacoralliens qui forment des charpentes rocheuses.
En conclusion, c’est bien la rapidité des changements de l’environnement marin qui est fatale à des communautés récifales fragiles. La conséquence sera un changement vers des communautés plus opportunistes : éponges, corail souple, macro-algues (voir figure 8 : un récif « dégradé » de Cayo Largo, Cuba). Ces organismes ne forment pas de charpente rocheuse et ne contribuent plus à la protection de l’érosion côtière.
La nature n’est pas la seule à en subir l’impact : dans un contexte de hausse du niveau de la mer de l’ordre de 3.3 mm par an actuellement, des millions d’habitants des îles coralliennes basses vont rapidement devenir des migrants climatiques.

Laisser un commentaire